
The general framework for developing a breeding programme is illustrated in Figure 2. The framework includes the previously discussed implications of agricultural policies, infrastructure and farmer involvement, markets and some aspects on the choice of populations available. A breeding programme needs to be integrated and its success is largely determined by the scope of farmer participation.
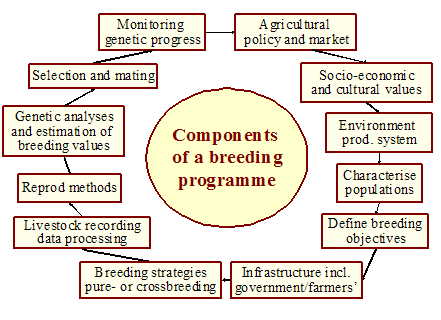
Figure 2. Components of a breeding programme.
In the following sections, different breeding strategies are presented and aspects on developing improvement programmes at breed level are dealt with. The scope of any breeding programme must be set in relation to the resources available and the stage of development in the region concerned. It must be kept simple and reliable, at least initially, rather than sophisticated and vulnerable to several prerequisites that cannot be guaranteed [CS 1.3 by Mpofu]; [CS 1.31 by Philipsson]. The design may, therefore, vary considerably depending on the actual breed, production system and other circumstances. Whatever the case, the principle of ‘KISS’ (keep it simple to be sustainable) should be emphasized in the start up phase of the breeding programme.
4.1 What strategy to choose
Payne and Hodges(1997) reviewed in detail the past developments of genetic improvement programmes for cattle in the tropics and what could be seen as the major options available for the future in seeking sustainable breeding systems. These differ for many reasons from the programmes designed for temperate breeds in Western countries. In summary, five major options are proposed:
- crossbreeding indigenous breeds with temperate breeds without AI
- improving indigenous breeds
- progressively substituting the breed with another indigenous breed
- crossbreeding indigenous breeds with temperate breeds using AI
- forming a composite(synthetic) breed.
The choice of strategy depends on many different factors specific to each situation and can be analysed according to the issues indicated by each one of the components illustrated. Generally, these options should aim at being simple enough to allow programmes to be launched without many resources. In most such cases, the schemes are based on open nucleus herds where the indigenous breeds are kept under selection. From these herds, males, either pure bred or crossbred, are distributed for use in smallholder or village farming systems. In this way, the indigenous breeds chosen for improvement will be conserved for the future. Depending on breed characteristics, the level of management and development one may choose a simpler or more advanced scheme. What have largely been neglected so far are options 2 and 3 in the bullets above. If comprehensive breed characterization is undertaken, it would be more likely to find interesting indigenous breeds for use far outside their present habitats—the success of the Sahiwal breed globally, the potential of the Kenana, Butana and N’Dama in Africa and the Nelore, Gir and Sindi breeds of Indian origin, but further developed in Brazil, deserve special attention in this respect [see Breed information]. Additionally, the additive genetic variation within the indigenous breeds seems to be large and has so far been little exploited [CS 1.36 by Sartika and Noor]. Formation of synthetic breeds has been tried and several breeds of today resulted from such practice [CS 1.26 by Ramsay et al.]; [CS 1.40 by Chako]. It is also an interesting way out of a situation when systematic crossbreeding does not work, while the incorporation of exotic genes is deemed important [CS 1.5 by Kahi]. However, the advantages of forming a synthetic breed from crossbreds must outweigh the loss of heterosis that otherwise could be achieved from systematic crossbreeding.
4.2 Defining the breeding objectives at breed level
The ultimate goals of a breed at themacro-level should be expressed by the agricultural development policy, market, production system and the output required from the resources available in the system of a country, region or locality [see FAO, 2010]. At the micro-level, the definition of breeding objectives means that the relative importance of improvement of different traits of the breed for a given production environment must be determined [Groen in ICAR Tech Series No. 3].In doing so, a long-term horizon of breeding should be kept. In cattle breeding, that means at least a time horizon of 10–15 years, while in pig and poultry breeding the considerably shorter generation intervals also allow for shorter time horizons in selection.
Breeding objectives must be set at the national, regional or local level by local stakeholders (and not by outsiders) to truly reflect the real needs of the area; farmers must support the direction of change (Ahuya et al., 2004; Kosgey et al., 2006; FAO, 2010) [CS 1.14 by Olivier]; [CS 1.28 by Madalena]. The conflicts that may occur between the long-term goals, expressed at national or organizational level and the interest of farmers in short-term benefits could be solved either by regulations or incentives for participation in a cooperative breeding programme.
When deciding upon a breeding strategy, though, the effects of longer time periods must be considered, consequently, when evaluating different crossbreeding strategies, several generations of selection and mating must be considered [Computer exercises: Breeding plans].
When determining the relative importance of different traits in the breeding objective one may, as an alternative to calculation of relative economic weights, put restrictions on the change in specific traits or define what the desired gain is in each trait. Whatever the choice of method of weighting traits, the following additional points must be considered:
- Although the long-term goals determine the breeding objectives and the role of each trait, the short-term benefits for farmers must be considered to get good farmer participation.
- In almost all situations, it may be difficult to exactly value the change in all desired traits in economic terms; fundamental traits must anyway be considered in the selection programme, e.g., through independent culling or other appropriate methods if the indexing procedure does not work or is not the appropriate approach.
- Special care must be taken in dealing with fitness and adaptive traits, especially if antagonistic genetic relationships exist between these and primary production traits.
The issue of whether to directly select in harsh environments for adaptive traits in addition to such important traits as production, reproductive performance and growth, is debatable. As physiological adaptability is expressed in performance, does selection of animals on the basis of performance alone give sufficient consideration to adaptive mechanisms involved in maintaining, say heat balance? Generally, favourable correlations suggest that adaptability traits would not be compromised by emphasizing selection for performance (Burrow et al., 1991; Mueller, 2006). There are, indeed, indications that selection for performance (e.g., reproduction, survival, growth etc.) in stressful environments will lead to selection for the most suitable animals (McDowell, 1972; Turner, 1984). Besides, as the number of traits in a selection programme increases, genetic progress that can be made in improving any one trait slows down unless the traits are highly genetically correlated [Computer exercises: Breeding plans]. Additionally, there are no satisfactory estimates of heritability of measures of adaptability in these populations. Moreover, cooperative breeding schemes, considered ideal for genetic improvement of indigenous livestock in developing countries, need to be as simple as possible initially and should, therefore, avoid complicated selection criteria (Kosgey et al., 2006). Nevertheless, one should, in most cases, try to focus selection on only the most important traits improving productivity and fitness for the environment in question [CS 1.19 by Yapi-Gnaore].
In a dynamic breeding programme seeking the optimal utilization of the genetic resources available, the breeding objectives should be reviewed regularly based on what has been achieved so far and on likely long-lasting changes of the market or agricultural policies [Groen in ICAR Tech Series No. 3]; [Weller in ICAR Tech Series No. 3]
4.3 Pure breeding or crossbreeding?
An early consideration, related to the choice of breeds, is whether the characterization of the indigenous breeds available shows that they have the potential for required improvement through pure breeding or if some kind of crossing with exotics or other tropically adapted breeds might be a better strategy.
The choice of breeding method, pure breeding alone, also using crossbreeding or breeding for a synthetic breed, is perhaps the most important decision to be made when designing a breeding programme. It relates partly to the previous discussion on characterization of genetic resources, including exotic germplasm being available on the international market. Key issues include:
- What are the level of performance and the potential of genetic improvement through selection within the indigenous breed?
- What alternative breeds are available for crossbreeding and what levels of performance and adaptability to the environment could be expected from 1st and 2nd generation crosses?
- How important are effects of heterosis for the traits of major interest?
- What are the opportunities for keeping pure bred stock of two or more breeds being available for maintaining a long-term crossbreeding programme?
- In the long run, what are the costs and benefits of crossbreeding compared to within-breed selection aimed at improving the same set of traits?
- Is the formation of a synthetic breed a viable alternative to both pure breeding and crossbreeding with other breeds?
Crossbreeding has principally been applied in the tropics to exploit breed complementarity. Specifically, specialized exotic (mainly temperate) breeds have been crossed with indigenous breeds to combine the high productivity of the former with adaptive attributes of the latter. Success stories are clearly available from countries that have good infrastructure (Madalena, 2005),[CS 1.5 by Kahi]; [CS 1.25 by Filho]; [CS 1.26 by Ramsay et al.]; [CS 1.28 by Madalena]; [CS 1.40 by Chacko]. However, many crossbreeding programmes have either lacked long-term strategies on how to maintain a suitable level of upgrading or have been too complicated to conduct in practice [CS 1.5 by Kahi]. As a result, uncontrolled crossbreeding has been identified as a major cause of loss of genetic diversity in indigenous breeds, primarily through replacement of pure indigenous breeds with crossbreds. However, these crossbreds may have provided the base for new synthetic breed developments, which thereby could carry on valuable genes from the original indigenous breeds to future generations.
On behalf of FAO, Cunningham and Syrstad(1987) extensively analysed results from crossbreeding in the tropics. Their clear conclusion was that consistent improvements in most performance traits were achieved in ‘upgrading’ dairy cattle to as much as 50% with temperate breeds. Beyond that, results were variable. Brazilian studies by Madalena et al. (1990a, 1990b) support these findings. Results vary according to environmental conditions and traits studied [CS 1.28 by Madalena]. A general conclusion is that crossbreeding to produce animals with up to 50% of the genes from temperate breeds can be recommended where crossbreeding is an option.
In practice, for cattle many schemes have been adopted where F1 heifers have been produced at government farms and been distributed before first calving to smallholder farmers. In this way, the farmers have then been able to raise their income through selling milk from improved cattle (Module 1, Section 2.3). However, as females are not replaced at the farm but mainly provided from government farms, this system is not sustainable and should only be utilized to introduce improved animals. The females should instead be produced through systematic crossbreeding at the village farm level. Brazilian experiences also show the opportunities for specialized production of pregnant F 1heifers for sale to specialized dairy farms. Results for dairy goats from FARM-Africa’s initiative in East Africa have demonstrated that this can successfully work (Ahuya et al., 2004) [CS Ojango et al]. However, the never-ending question has been what breed to use when mating the F1 animals. Usually schemes that are too complicated have been proposed to maximize the genetic gain, considering both additive and non-additive genetic effects. Too often the programme has failed due to the practical difficulties of running the scheme if sufficient infrastructure is not in place.
It is important, therefore, to find a simple crossbreeding strategy which can easily be followed under practical conditions. For simplicity and cost reasons (Peacock et al., 2011), it should be based on continuous use of the females produced in the herd, allowing new males to enter the herd live or by AI. Figure 3 exemplifies such a plan. It is based on continuous use of F1 males on indigenous females and by time on crossbred females, in village herds and allows a maximum of 50% exotic genes to be incorporated in the female stock. The strategy is based on two cornerstones:
- A nucleus herd of selected animals of the pure indigenous breed is kept for continuous selection within the breed and for mating with an exotic breed to produce F1 males for distribution to village herds.
- Crossbred females in the village herds are bred to new F1 males from the nucleus herd to produce the next generation of females at farm level.
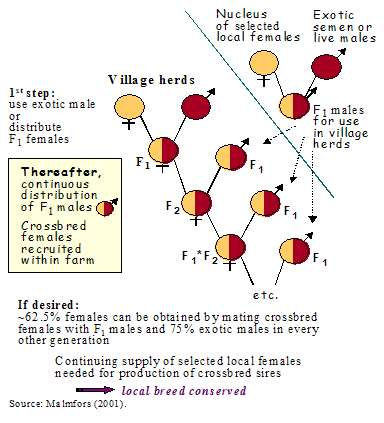
Figure 3.Example of crossbreeding scheme for livestock.
This strategy leads to animals that on average contain 50% of the genes from the indigenous breed and 50% from the exotic breed. To speed up the programme, F1 females can be produced directly by using exotic males, or semen from these, for mating the females in the village herds. If a higher degree of upgrading is desired, e.g., 60–65%, then the nucleus herd should produce males that initially have 75% exotic genes, but later also F1 males for rotational use. The 50% plan is quite simple, while the other one starts to become a little complicated. If the nucleus herd for some reason fails to produce the F1 males, there is an opportunity to continue to select both females and males within the herd populations established at village level. A synthetic breed or population is then underway. However, the degree of success will depend on the extent to which villagers are involved in the design, implementation and review of stages of the performance and pedigree recording system (Okeyo, 1997; Ahuya et al., 2004, 2005; Kosgey et al., 2006; Peacock et al., 2011)
The advantages of this crossbreeding scheme are:
- crossbred females are recruited within the village farms
- only crossbred males need to be distributed
- no risk of too high‘upgrading’
- simple method, requiring minimum infrastructure
- local breed will be conserved (in nucleus herd), genetically superior individual indigenous animals resulting from each generation of selection can be used to improve a wider population of the indigenous breed in production systems and environments where they are best suited. If the programme for any reason fails it will not lead to erosion of the initial genetic resources, but as an alternative, maintain these in a new synthetic population.
The disadvantages are:
- Heterosis is not maximized (but the complementarity of additively inherited traits is exploited, i.e., desired traits are selected for and combined).
- Some segregation in crossbred females may occur but this could be counteracted through selection within the village herds.
Overall, it is believed that the advantages of this simple scheme by far outweigh the disadvantages mentioned. However, inter se mating of F1animals to produce F2animals are less predictable in performance than F1animals, partly dependent on breed combination and its interaction with the environmental conditions. It is therefore advisable to research, or at least carefully monitor, the results of each generation before wide-scale implementation of the proposed scheme.
To improve dairy cattle, it seems often obvious to apply crossbreeding with temperate breeds in tropical farming systems. However, some indigenous breeds such as the Kenana in Sudan and the Sahiwal in both Asia and Kenya have potentials to be used in pure breeding and for crossbreeding with other indigenous Bos indicus breeds [CS 1.31 by Philipsson]. The Jamnapari goats of India would play similar roles in improving milk and meat production in goats under tropical conditions. For beef production, several high potential indigenous breeds are available for tropical environments, e.g., the Boran cattle in Ethiopia and Kenya, Brahman in India, Tuli, and Nguni breeds in Zimbabwe and South Africa respectively and Nelore in Brazil.
The choice of breeding strategy, i.e., pure vs. crossbreeding has vast implications at both farm and organizational levels and should be based on facts that are as extensive as possible [FAO, 2010]. Any change in the direction towards crossbreeding from an ongoing pure breeding strategy should be preceded by research that provides information on performance, reproduction and health, including adaptive traits [CS 1.23 by Fall]. Such information is needed at least for the F1 animals and their backcrosses to the exotic breed compared with pure breds in representative environments. Serious attention must also be paid to the logistic aspects of organizing the crossbreeding programme to be sustainable.
4.4 What type of livestock recording schemes and data processing may be available?
The goal of livestock recording schemes is usually to provide farmers with information about individual animals for management and for breeding purposes. The objective could also be phrased as to provide aninformation system about the livestock, their use, performance and development, by both farmers and national authorities. The available infrastructure, including physical and human resources, will determine the type of recording scheme that can be effectively implemented. The sort of scheme offered will differ considerably depending on the farming structure and production system. Early stages of development require simple solutions to be sustainable. With time and experience, the schemes may be made more sophisticated (Kosgey et al., 2011). It is better to start recording in a few cooperating herds that can be handled well, rather than running a scheme on a wide scale that cannot be supervised efficiently. In any new recording scheme efforts should be made to incorporate, where possible, the existing indigenous systems and institutions to ensure quick adoption and success.
Nucleus herd breeding schemeswhere the selection of breeding stock is concentrated in a few herds from which the selected animals are spread to other herds, are attractive in many developing countries as suggested by Smith (1988). They are designed to allow a good recording on a limited number of animals and data management at reasonable cost, and may be combined with the use of efficient reproduction technologies [CS 1.7 by Khombe]. Open nucleus breeding schemes, which also allow inflow of high potential breeding animals from other herds, have been proposed as ideal for genetic improvement in situations with moderate levels of management (Smith, 1988; Barker, 1992). A nucleus herd programme is used to both conserve an indigenous breed and to upgrade the local population (see Figure 4).

Figure 4.Open nucleus herd breeding scheme—basis for conserving an indigenous breed and upgrading local population.
As recording schemes include different activities and serve various purposes but involve the same farmers and animals, the activities must be well integrated to be cost-effective and provide the farmer with added information [CS 1.13 by Banga]; [CS 1.14 by Olivier]; [CS1.19 by Yapi-Gnaore]; [CS 1.26 by Ramsay et al.]. There are many examples around the world where good attempts at recording different aspects of the livestock partly lose their value as the information is not integrated and fully utilized (Wasike et al., 2011); [CS 1.19 by Yapi-Gnaore]; [ICAR Tech. Series No. 1. 1998]. Alternative forms of dispersed nucleus breeding schemes and modified sire reference schemes have been successfully implemented in Latin America and parts of Asia (Mueller, 2006).
4.5 What reproduction technologies are feasible?
AI has undoubtedly proven its value for genetic improvement programmes of several species, but most notably in cattle. The success in cattle breeding depends on the number of pregnancies it is possible to achieve per bull and per year through AI compared to males of other species. The utility of frozen semen in cattle is also exceptionally good. These advantages have also proven beneficial in many developing countries, not the least in crossbreeding when genes of exotic breeds have been introduced through semen imports. The widespread use of AI in the Kenya highlands of East Africa, for example, provided the opportunity of introducing milk for school lunches in the country as a result of increased production of milk from crossbreeds and exotic breeds. This technology has also lead to one of the most successful smallholder dairy systems in the developing world apart from the Indian example (Stall et al., 2008a). However, with the sudden change of policy and removal of public support, the system simply collapsed. The development of Sunandini cattle as a synthetic breed through consistent use of AI provided almost a tenfold increase in the per capita consumption of milk among the people of Kerala Province in southern India [CS 1.40 by Chacko]. Therefore AI has great advantages from a genetic improvement point of view through its effective dissemination of germplasm and the opportunities for strong selection of breeding stock [CS 1.31 by Philipsson]. Equally important, this methodology has great advantages in controlling or eradicating diseases that might be transmitted in natural mating systems.
However, the use of AI has also failed in many situations in developing countries because of the lack of infrastructure and the costs involved, such as for transportation and liquid nitrogen for storage of semen or because the breeding programme has not been designed to be sustainable[CS 1.3 by Mpofu]; [CS 1.31 by Philipsson]. Improper use of AI for crossbreeding indigenous cattle with exotics may be disastrous when, for example, a long-term strategy lacks information on how to maintain the appropriate level of exotic genes in an environment that cannot support pure exotic breeds. The pros and cons of using AI should therefore be critically reviewed for each case before designing breeding programmes
Another reproduction technology proven to enhance genetic progress in many situations is embryo transfer (ET). Superior females may be super-ovulated and mated with highly selected males to produce embryos of high expected genetic merit. Such systems suit nucleus-breeding schemes well and provide specific opportunities for conservation and development of minor breeds, for establishment of gene banks and for synthetic breed formation. Provided the technique and infrastructure are available, it may also be useful in developing countries [CS 1.16 by Mpofu]. However, the costs versus benefits must be critically evaluated, considering actually obtained and not ideal technical results. Use of sexed semen, either alone or via in vitro production have also been tried, but no commercial success has been achieved to date. Scenarios where use of sexed semen in combination with genomic selection has been simulated by Pedersen et al. (2010), however, the results showed non-significant potential genetic gains, and even fewer gains are possible when such strategies are used in combination with multiple ovulation and embryo transfer (MOET) technology.
4.6 What methodologies for genetic evaluations should be applied?
Tropical regions are endowed with a wide diversity of breeds and strains of livestock. Besides, the available body of evidence indicates that there is substantial within-breed variation in most of the economically important traits. Indeed, estimates of heritability of these traits in tropical breeds in well-managed populations are often either within the range of or higher than corresponding estimates from temperate regions. Since most populations of indigenous tropical livestock have been subjected to only very mild artificial selection pressures for productivity, the general trend of high heritability estimates is expected [CS 1.6 by Mpofu and Rege]; [CS 1.9 by Aboagye]. However, the few available estimates of heritability for production traits in indigenous tropical breeds have invariably been based on insufficient data. Furthermore, most of these studies have suffered from poor experimental design. These factors, and the generally poor animal management in these situations, have obviously resulted in large environmental variations and biases. Therefore, heritability estimates on the lower end of the scale have often resulted from large environmental variation rather than from small genetic variation. The low reproductive performance of tropical cattle may largely be due to environmental, mainly nutritional, stresses. Nonetheless, estimates of heritability of female fertility traits in the tropics, while low to moderate, are usually higher than estimates in temperate cattle breeds.
Another critical component of genetic improvement, apart from ‘variation’ is selection intensity. In low-input (traditional) systems, reproductive rates are often so low, especially in cattle, and mortalities so high that there is hardly any opportunity for selection. Farmers invariably have to keep all female animals that survive, not because they are they most productive, but because they are hardy. Absence of recording is another serious constraint, which makes it impossible to undertake selection on objective criteria. Selection pressure is further compromised in most cases by small herd sizes and uncontrolled breeding in communal grazing systems (Rege et al., 2011).
The basic principles for genetic evaluations based on pedigrees, individual performance and sib and progeny information are, however, always valid. Generally, the more information included from the individual and its close relatives, the more accurate the estimated breeding values will be. However, three points need to be made:
- The concept of progeny testing as a method for genetic evaluation and selection is widely over-emphasized, especially if the breeding programme does not allow a rather high level of infrastructure and sophistication and if the populations are small.
- Because of the large environmental influences on production in many tropical production systems, it is tempting to use rather advanced genetic statistical methods of analysis of the performance data, i.e., BLUP (Best Linear Unbiased Prediction) Animal Model, to correctly separate genetic and environmental effects and to consider all genetic relationships [Group discussion: Breeding programmes. However, using BLUP does not comply with the KISS principle! It may therefore be advisable, at least at the beginning of a selection scheme, to select just on phenotypic values within similarly raised or kept animals of the same age. With time, more advanced methods for genetic evaluation may be applied. Current progress in molecular genetics indicates that information on genetic markers associated with specific traits, e.g., disease resistance and quality of products, may become useful in the future (and would be cheaper) as a complement in genetic evaluations, as is indicated in the next section [CS 1.19 by Yapi-Gnaore].
The use of mass selection (animals selected on phenotype), including pedigree information, seems to provide the best base in many situations for correct ranking of potential breeding stock in developing countries, especially for animals held in nucleus herds with good record keeping. Mass selection is also a valuable method for screening animals to form the initial nucleus population. Animal identification systems that use already existing indigenous traditional knowledge and simpler methods such as scoring and ranking of only the top 5–10% of animals in the herd, where herds are large as in pastoral communities, would provide a good avenue for using more accurate genetic evaluation methods. Within traditional livestock production systems livestock keepers (e.g., pastoralists) can identify and rank their stock accurately. Ranking methods used within these systems can be documented and practically applied if the livestock keepers are involved in the design of evaluation programmes from the outset (Kosgey and Okeyo, 2007).
4.7 DNA-analyses and genetic marker information useful for selection and introgression
The hitherto most successful proven use of molecular genetics in practical breeding programmes relates to identification of single genes which in their recessive homozygous forms are lethal or bring defects to the animal. Successful DNA-tests have been developed for a number of such genetic defects, e.g., Bovine Leukocyte Adhesion Deficiency (BLAD) and Complex Vertebral Malformation (CVM) in cattle, which enable their elimination from the populations in question. BLAD, an immune deficiency, and CVM, a vertebra malformation, are defects widespread in the Holstein cattle breed caused by recessive genes. In both cases, carrier bulls had been used widely around the world before the defects were discovered [See OMIA] or the website (http://omia.angis.org.au/).
Current progress in molecular genetics has also shown that information on the genetic background of quantitative traits will be available to an increasing extent in livestock. Recent developments, following the sequencing of whole genomes of several species, have opened up completely new opportunities. By correlating the DNA information, single-nuclear-peptides (SNP), in the chromosomes with phenotypic information of the same animals it is possible to develop regression formulas for prediction of breeding values of future animals. Thus, breeding values can be predicted for the newborn animal with an accuracy almost twice as high as that obtained with just pedigree information. The gain is larger for traits with low heritability than for those with high heritability. Practical use of the methods began in 2009 in dairy cattle breeding, but it is still difficult to evaluate the efficiency of the methods practised (Reents, 2010; Wiggans et al., 2010; Weigel., et al., 2010). Four important pre-requisites must be mentioned:
- In order to develop the regression formulas to predict breeding values of future animals it is necessary to base these calculations on large sets of population data on phenotypic information of all traits for which breeding values are going to be predicted. So far, when using panels of 50K SNPs in dairy cattle several thousands of bulls with accurately estimated breeding values have been needed in the so-called reference or training populations which contain both SNP information and breeding values based on phenotypic records.
- Due to selection based on SNP information the prediction formulas need to be regularly revised. Thus, continued phenotypic recording of important traits is still needed.
- Results for one breed are not directly applicable in another breed, i.e., data and prediction formulas need to be developed for each breed where the methodology is going to be practised.
- Although costs for genotyping individual animals are rather high they will most likely reduce, but will still be high for large-scale use.
A specific area of use of genomic evaluation is for parentage testing. Also, in cases where no pedigree information is available it may be possible to determine the genetic relationships among animals in a population, provided enough SNPs are available for the analysis.
The area of genomic evaluation and selection is under very rapid development, not least the technical tools to use. The use of SNP panels of variable size will be available, e.g., panels with 3K for screening purposes as well as 800 SNPs for more accurate predictions in dairy cattle. Also whole genome evaluations are in the pipeline (Interbull Bulletin 41, 2010). Whatever happens, no progress in the use of genomic selection is possible without accurate phenotypic records of large populations. That is why it is still urgent to develop livestock recording in the developing world.
Previous research and use of molecular genetic information has focused on identification of quantitative trait loci (QTL) for economically important traits in many different species and breeds. Thus in some cases, information on QTL (Module 4, Section 6)added valuable information to breeding programmes by so-called marker-assisted selection (MAS). Its greatest impact in practical situations was expected to be where phenotypic information is limited, e.g., when the heritability of the traits in the breeding goal is low, when the trait is expensive to record, when the traits cannot be recorded on all individuals (sex-limited traits, carcass traits and disease traits).
When QTL has been located accurately, marker information can be used in the genetic evaluation of individuals even if the gene itself is not pinpointed. Potentially, MAS can increase genetic progress by increasing accuracy of evaluations, by increasing selection intensities and by decreasing generation intervals (Dekkers, 2004; Cole et al., 2010). For dairy cattle, for example, several different breeding schemes using MAS have been proposed (reviewed by Weller, 2001; Meuwissen, 2005). A recent review by Marshall et al. (2011) and results of a study by Bennett et al. (2010), show that unlike gene assisted selection (GAS), MAS could be of limited application in some traits, especially in dairy and beef cattle. However, MAS has potential application in poultry, pigs and beef cattle breeding. In progeny testing schemes marker information can be used in addition to phenotypic information from the daughters of sires and for pre-selection of young bulls before entering progeny test.
These advantages are even more obvious with genomic selection as this method makes use of not only QTLs but also all small genetic effects of the other loci in the genome.
Introgressionis a breeding strategy for transferring specific favourable alleles from a donor population to a recipient population. This would, for example, be of great interest for genes responsible for disease resistance, which could be introgressed into a susceptible but otherwise economically superior breed. The strategy has two components:
- fixation of the favourable alleles in the recipient population
- reduction or elimination of the rest of the donor genome from the recipient population.
Crossing the donor and recipient population produces an F1 generation. Thereafter, a series of backcrosses to the recipient population is performed, but in each generation only individuals that carry the favourable donor allele are selected to produce the next BC generation. After a number of back crossings, the progeny are inter-crossed and a population that is homozygous for the donor allele is obtained. The higher number of BC generations before the inter-cross and the larger proportion of the genome will then be from the recipient population. When the gene to introgress is a QTL, genetic markers must be used to identify the favourable donor allele. Markers can also be used to identify the origin of the remaining genome and therefore decrease the number of BC generations needed. The DNA-based marker identification technology is becoming cheaper with time thus enabling MAI to become affordable, even under low-input production systems, so long as the breeding programme is effectively organized. Such a MAI scheme is shown in Figure 5.
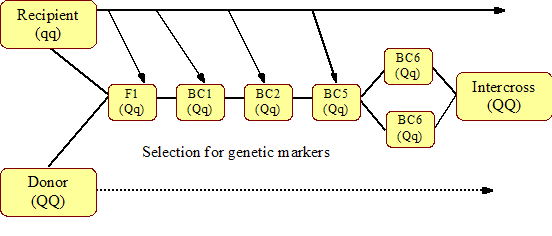
Under tropical conditions, there are presently two obvious candidate traits for MAS and MAI-trypanotolerance in African cattle and helminth resistance QTL in sheep. Several QTL related to trypanotolerance have been detected in an experimental cross between the trypanotolerant breed N’Dama (B. taurus) and the East African Boran (B. indicus) as reviewed by van der Waaij (2001), while helminth tolerance QTL in sheep is currently being studied at ILRI in experimental crosses between the helminth tolerant Red Maasai breed and the susceptible Dorper sheep breed. These QTL, if found to be of substantial influence, could be used for MAI, MAS or GAS if the associated genomic area/SNPS were to be identified; they could also be used in a combined introgression and selection programme. A drawback with a pure MAI programme is the large number of individuals required, and ensuing high costs, when several QTL are to be introgressed. There is also a possibility that other advantageous genes besides the mapped QTL or positive gene combinations are lost when the donor genome is eliminated. This loss of genes/gene combinations is also a risk when only marker information is used for selection within a breed or in a hybrid population; use of MAI and MAS in addition to conventional methods is therefore recommended. However, an important advantage with QTL information for resistance to diseases is that animals can be selected without exposing them to infection. That is, individuals or embryos that carry the required genes can be selected as early as immediately after conception. The individuals or embryos can be further tested for the other desirable genes for high growth, carcass quality etc. soon after birth.